
Der Ursprung der Muskelkraft? Vom Sarkomer zum Muskelbündel
Stand: 2019
So wie die Ausdauerfähigkeit mit dem Herz-Kreislauf-System in Verbindung steht, so ist die Kraftfähigkeit untrennbar mit der Muskulatur verbunden. Auch wenn die unterschiedlichen Funktionssysteme im menschlichen Körper nicht getrennt voneinander betrachtet werden können, kommt der Muskulatur bzw. dem neuromuskulären System eine besondere Bedeutung in diesem Zusammenhang zu. Trainer sollten dabei eine Vorstellung haben, wie die Muskulatur grundsätzlich aufgebaut ist und welche Prozesse im Körper ablaufen, bevor eine Kraftwirkung nach außen sichtbar wird.
Für Trainer ist die willentlich steuerbare Skelettmuskulatur von besonderer Bedeutung, da sie den eigentlichen motorischen Aktivitäten dient. Wie es der Name schon sagt, sind unsere in der Regel 501 Skelettmuskeln 1 dabei meist über Sehnen mit Skelettknochen verbunden, die dabei Widerlager und Hebel für die Krafterzeugung darstellen. Der innere Hebelarm als Kraftarm besteht i. d. R. aus einem Knochenvorsprung und fällt meist sehr viel kürzer als der Lastarm aus. Dadurch müssen die Muskeln in der Regel sehr viel größere Kräfte aufwenden, als man von außen vermuten mag. Zum waagrechten Halten eines 1 kg schweren Balls muss z. B., vereinfacht ausgedrückt, unser Biceps brachii statt ca. 10 N in etwa das 10-fache, also 100 N erzeugen. Hier liegt eine anatomische Eignung für eine Sportart oder Disziplin begründet.
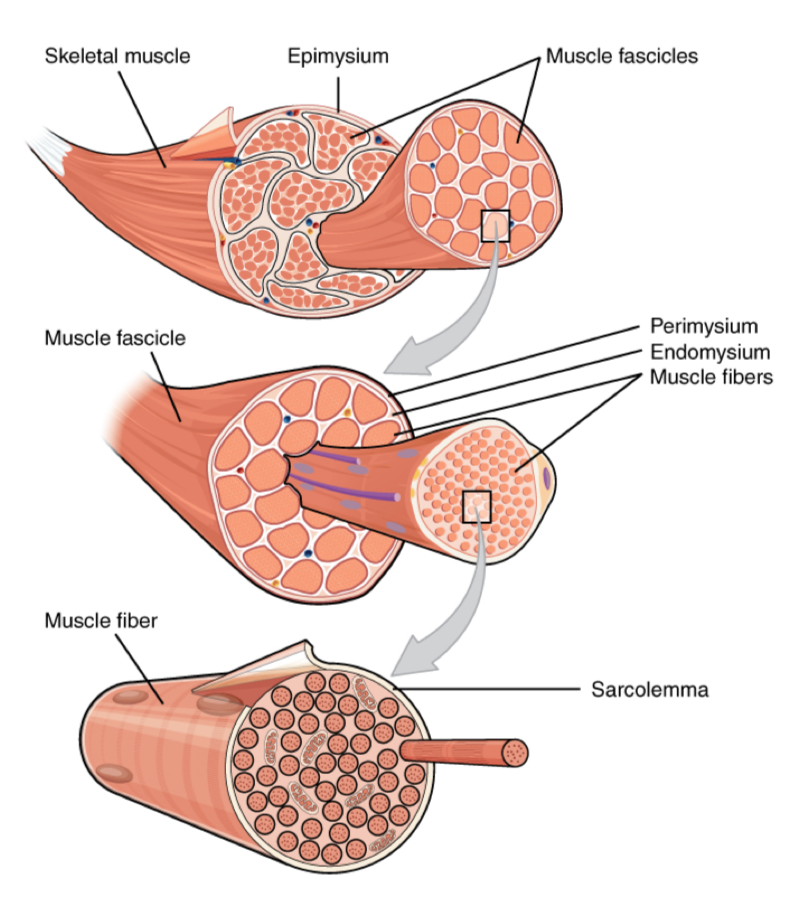
Abb. 1: Aufbau der Skelettmuskulatur 4
Grundsätzlich ist jeder Skelettmuskel strukturell hierarchisch aufgebaut (Abb. 1). Ein Skelettmuskel besteht in der Regel aus mehreren Muskelfaserbündeln (Faszikeln), die wiederum aus zahlreichen Muskelfasern, den eigentlichen Muskelzellen (den sog. Myozyten) bestehen. Diese Myozyten sind vielkernige, einige Mikrometer dicke und mit bis zu 40 cm sehr lange Zellen, die jeweils ca. 1000-2000 sog. Myofibrillen enthalten 1 , 2. Jene Myofibrillen stellen das zentrale strukturelle Merkmal von (Skelett-)Muskelzellen dar und verlaufen wie „Fäden“ parallel in der Zelle. Sie sind es, die eine willentliche Verkürzung oder Entspannung des Muskels ermöglichen. Dabei sind Myofibrillen ihrerseits aus jeweils mehreren hundert „in Reihe“ geschalteter sog. Sarkomere aufgebaut. Jene einzeln ca. 2 μm langen Sarkomere stellen die eigentlichen kontraktilen Funktionselemente des Muskels dar 1 , 2. Vielfach parallel aneinandergereiht, geben sie einer Muskelfaser im Lichtmikroskop ein charakteristisches quergestreiftes Erscheinungsbild, was der Skelettmuskulatur ihren Beinamen gegeben hat 2 . In den Sarkomeren, die an den sog. Z-Linien (Zwischenlinien) miteinander verbunden sind, agieren drei wichtige Muskelproteine: Das dünne filamentartige Strukturprotein Actin, das dicke Motorprotein Myosinfilament (bzw. eine ganze Familie dieser Proteine) sowie das filamentartige Titin. Vereinfachtausgedrückt verbindet ein Myosin-Protein zwei Actin-Filamente, die wiederum durch je ein Titin-Filament mit den angrenzenden Z-Scheiben und damit den benachbarten Sarkomeren verbunden sind 1 , 2 , 3.
Ein willentlicher Aktivierungsreiz wird über spezielle Nerven, die sog. Motoneuronen, an die Muskelfaser übermittelt. Über die sog. motorischen Endplatten, welche sich an den Enden der Motorneuronen befinden, werden elektro- und biochemische Prozesse in der Muskelfaser eingeleitet. Vereinfacht ausgedrückt, bewirkt ein elektrischer Nervenimpuls die Freisetzung von Kalziumionen, die wie ein „Schlossöffner“ auf die Actin-Myosin-Verbindungen wirken, sodass das Myosinmolekül mit seinen sog. Köpfchen radial an seinen beiden Enden Schritt für Schritt in jeweilsentgegengesetzter Richtung an den Actin-Filamenten „entlangwandern“ kann 1 . Für jeden einzelnen Schritt muss dazu Energie aufgewandt werden, die biochemisch durch die Umwandlung von Adenosintriphoshphat (ATP) zu Adenosindiphosphat (ADP) sowie einem Phosphatmolekül frei wird 1 , 2. Durch das mit seinen beiden Köpfchen an den benachbarten Actinfilamenten entlangwandernde Myosinmolekül erhöht sich der Überlapp mit den Actinfäden, ähnlich wie die Finger von zwei Händen, die dichter ineinander gleiten. Das Sarkomer verkürzt sich dadurch in Längsrichtung, es kontrahiert. Nach jedem Wanderungsschritt wird die eingesetzte Energie aus der ATP-ADP-Umwandlung als Wärme an das umliegende Gewebe abgegeben – der Grund, weswegen durch Muskelarbeit stets spürbar Wärme erzeugt wird 1 , 2. Hört die nervale Aktivierung auf, können die Myosinköpfchen wieder in ihre Ausgangslage zurückgleiten, wodurch sich die Sarkomerlänge wieder verlängert. Erst seit Kurzem weiß man übrigens, dass nicht nur die Actin-Myosin-Verbindungen, sondern auch das elastische Titin aktiv zur Sarkomerkontraktion beiträgt 3.
Da die Aktivierung aller Sarkomere einer Muskelfaser grundsätzlich über dieselbe motorische Endplatte und somit über dieselbe „Telefonleitung“ erfolgt, gilt für die Muskelfasern des Skelettmuskels das „Alles oder nichts“- Prinzip. Das heißt, alle Myofibrillen einer Muskelfaser können nur gleichzeitig kontrahieren oder entspannen, wodurch die gesamte Muskelzelle entweder maximal oder gar nicht kontrahiert 1 , 2. Grundsätzlich werden alle willentlichen Kontraktionen unserer Skelettmuskeln nicht durch einzelne elektrische Nervenimpulse, sondern durch eine Vielzahl kurz hintereinander ausgesandter Impulse gesteuert, man spricht dabei auch von Frequenzierung 1 , 2. Durch diese kurze Abfolge elektrischer Kontraktionsreize überlagern sich mehrere Einzelkontraktionen der Muskelzelle, wodurch die Gesamtkraft einer Muskelfaser im Vergleich zu einer Einzelkontraktion mehr als verdoppelt wird2 . Bei dieser Form der frequenzierten nervalen Muskelaktivierung spricht man auch von (vollständiger) tetanischer Kontraktion (Tetanisierung). Einzelzuckungen treten bei gesunden Skelettmuskeln hingegen nur bei Eigenreflexen und somit unwillkürlich auf 1 , 2.
Wie ist es nun aber trotz dieses „Alles oder nichts“-Prinzips möglich, dass wir sehr wohl die Krafterzeugung z. B. unserer Gliedmaßen oder Hände sehr fein abstufen können? Hier macht sich die Natur das Prinzip der varia- blen Rekrutierung zu Nutze: Zwar kontrahieren nach obigen Überlegungen einzelne Muskelfasern (tetanisch) entweder zu 100 % oder 0 %, doch können durch die (uns kognitiv nicht bewusste) Wahl, welche Muskelfasern eines Muskels kontrahieren, entweder nur wenige oder zahlreiche Muskelfasern eines Muskels gleichzeitig aktiviert, man sagt auch rekrutiert, werden. Eine Gruppe von mehreren Muskelzellen, eine sog. motorische Einheit, wird dabei jeweils von einem separaten Motoneuron innerviert. Je nachdem, welche und wie viele Motoneuronen gleichzeitig „feuern“, kontrahieren wenige oder zahlreiche motorische Einheiten des Muskels. 1 , 2. Unser Gehirn und Rückenmark leisten dabei eine beachtliche logistische Arbeit, indem sie für diese willentliche Kraftentwicklung i. d. R. ständig andere motorische Einheiten rekrutieren, wodurch eine Art „Rotation“ zwischen den Muskelfasern wie bei einer (sehr großen) Fußballmannschaft stattfindet. Auf diese Weise wird eine einzelne Muskelfaser effizient vor energetischer und struktureller Überlastung geschützt 2 . Erst dann, wenn die aufzuwendende Kraftanstrengung maximal wird, muss der Großteil aller motorischen Einheiten eines Muskels gleichzeitig rekrutiert werden, wodurch dieser dann jedoch auch schnell ermüdet. In diesem Ausnahmezustand einer Maximalanstrengung kann man das Zusammenspiel von hoher Frequenzierung und synchroner Rekrutierung des Großteils aller Muskelfasern eines Muskels, man spricht auch von Synchronisation, gut beobachten: Der Muskel beginnt sichtbar zu „zittern“ – ein deutliches Zeichen, dass die Intensität der Krafterzeugung nicht mehr lange aufrechterhalten werden kann 2 . Für den Sportler ist überdies interessant zu wissen, dass die Fähigkeit, möglichst viele Muskelfasern gleichzeitig in einem Muskel zu rekrutieren, auch vom Trainingszustand abhängt. Während ein untrainierter Mensch willentlich nur ca. 50 % aller Muskelfasern eines Muskels gleichzeitig rekrutieren kann, erreicht ein trainierter Athlet ein Verhältnis von bis zu etwa 90 %. Die letzten Prozentpunkte der Synchronisation sind unter normalen Bedingungen willentlich nicht erreichbar, sondern fallen unter die sog. autonomen Reserven. Diese sind nur unter psycho-physiologischen Extremzuständen, wie z. B. Todesangst, Drogen bzw. Doping, auszuschöpfen 1.
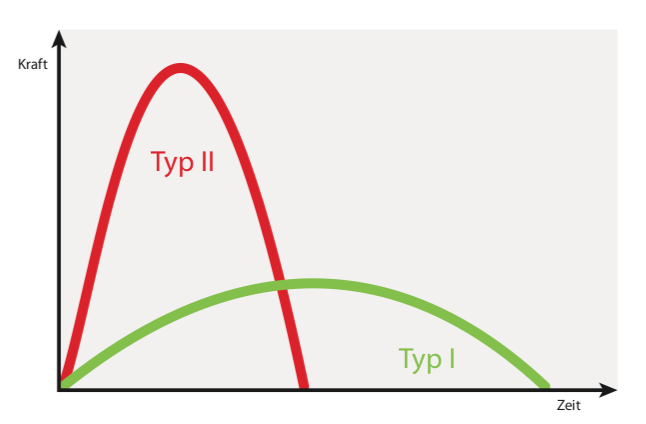
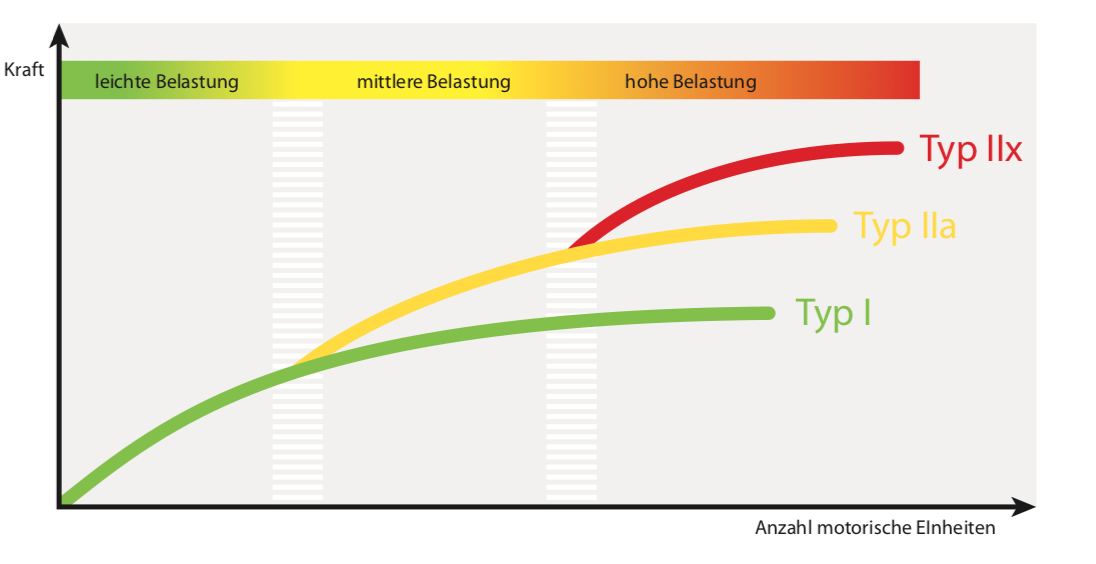
Abb. 2: Rekrutierungsfolge nach Belastungsintensität (verändert nach2)
Unabhängig von der Art und Weise der neuromuskulären Ansteuerung gibt es auch hinsichtlich der Muskelzellspezialisierung Unterschiede zwischen verschiedenen Muskelfasern von Skelettmuskeln. Grundsätzlich unterscheidet man nach moderner Nomenklatur drei histologische Grundtypen von Skelettmuskelzellen: Den langsam kontrahierenden Typ I („langsam zuckend“, engl. „slow twitch“, ST-Fasern) sowie die schnell kontrahierenden Typen IIa und IIx („schnell zuckend“, engl. fast twitch, FT-Fasern). Während die ST-Fasern (Typ I) nur vergleichsweise geringe Kräfte erzeugen, können sie ein konstantes Kraftniveau über eine lange Zeit aufrechterhalten. Die FT-Fasern (Typ IIx) hingegen können vergleichsweise große Kräfte erzeugen, jedoch ermüden sie schnell 1 , 2. Diese unterschiedlichen Eigenschaften resultieren aus einem histologisch unterschiedlichen Aufbau (Konzentration von Myoglobin, Anzahl an Mitochondrien etc.), der sich auch in differierenden primären Energiebereitstellungsmechanismen (oxidativ oder glykolytisch) ausdrückt. Abhängig von der zu erzeugenden Kraft rekrutiert unser Körper im Allgemeinen zuerst die Typ-I-Fasern eines Muskels, erst dann die Typ-IIa- und nur bei großer Kraftanstrengung auch die Typ-IIx-Fasern (Abb. 2). Diese gestaffelte „Zuschaltung“ wird auch als „Henneman’sches Rekrutierungsprinzip“ bezeichnet 2 .
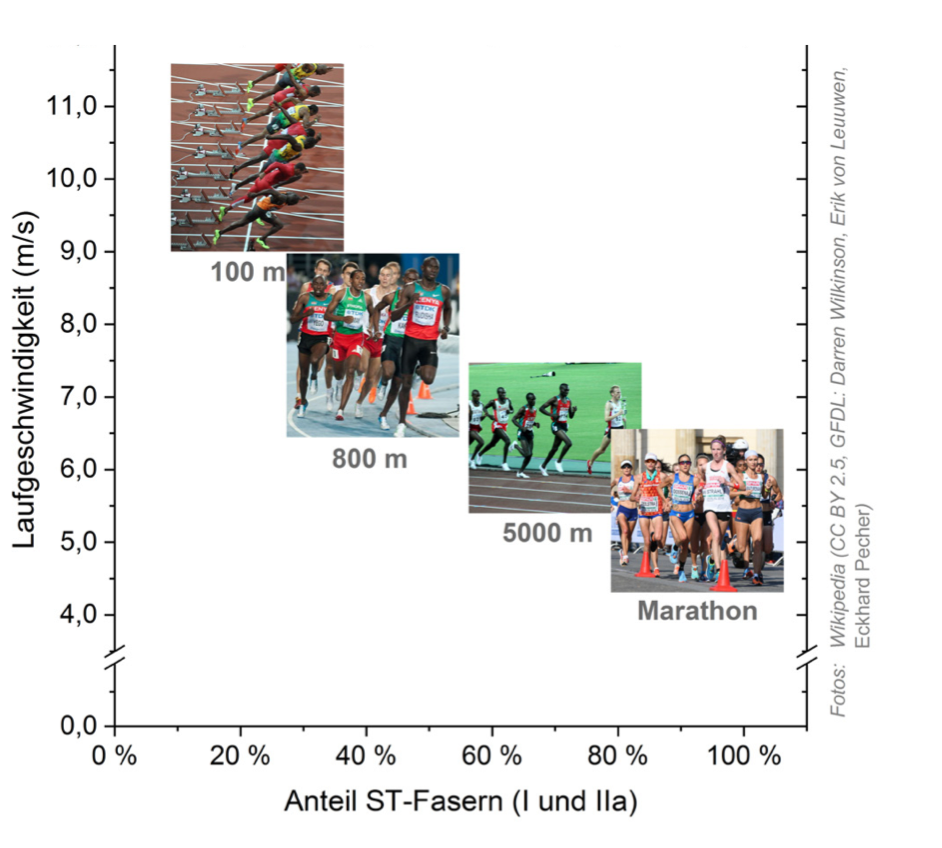
Abb. 3: Muskelfaserspektrum in verschiedenen Laufdisziplinen (verändert nach 2)
Abhängig davon, welche Kraftform (z. B. explosiv oder ausdauernd) für eine Sportart primär erforderlich ist, findet man in den erfolgreichen Athleten verschiedener Disziplinen ein unterschiedliches Muskelfaserspektrum. So haben die weltbesten 100-m-Sprinter beispielsweise einen sehr hohen Anteil an FT-Fasern im Körper, der über 70 % liegen kann 2 (Abb. 3). Spitzenmarathonläufer verfügen hingegen nur über einen geringen Anteil an FT-Fasern und besitzen stattdessen zu über 80 % ST-Fasern 2. Für die Normalbevölkerung liegt der Anteil an ST-Fasern bei etwa 50-60 %, der von FT-Fasern entsprechend bei 40-50 %, wobei individuell große Varianzen bestehen 2.
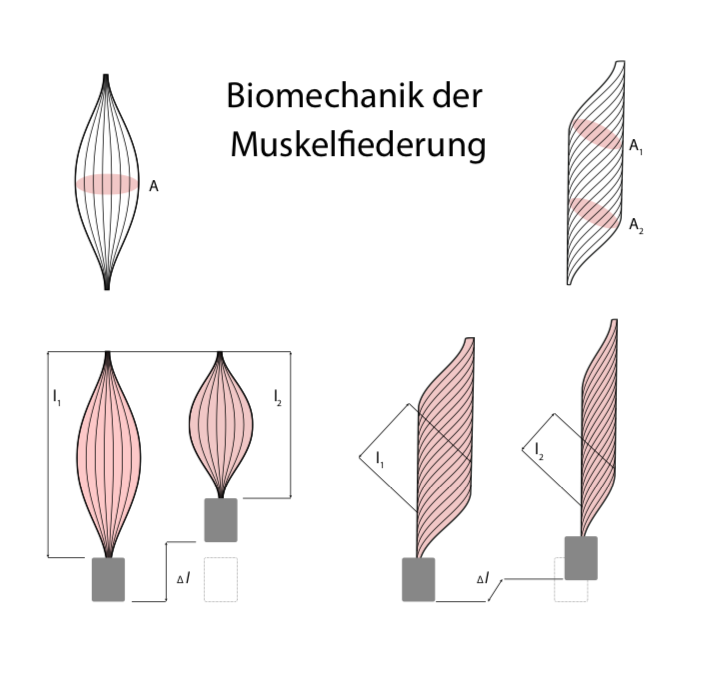
Abb. 4: Spindelförmige und gefiederte Muskeln (verändert nach 1) .
Unabhängig von diesen „Bauplanunterschieden“ auf zellulärer Ebene gibt es auch einen strukturellen Unterschied auf Muskelbündelebene, der erheblich über Kraft- und/oder Schnelligkeitsfähigkeit eines Muskels entscheidet: die sog. Fiederung. Verlaufen die Muskelfasern nicht parallel zum äußeren Muskelverlauf als Muskelspindel, sondern bilden einen sog. Fiederungswinkel mit ihm, unterscheidet sich der für die Muskelkraftentwicklung tatsächlich wirksame sog. physiologische Muskelquerschnitt von dem anatomischen Muskelquerschnitt (Abb. 4). Dies bewirkt, dass bei gleichem äußeren (d. h. anatomischen) Muskelquerschnitt deutlich mehr Muskelfasern kontrahieren und gemeinschaftlich „an einem Strang ziehen“ können als im Fall des spindelförmigen Muskels. Die Natur hat hier also einen eleganten Weg gefunden, auf begrenztem „Bauraum“ bzw. unter Gewichtsoptimierung mehr Muskelfasern unterzubringen: Bei gleicher äußerer Größe bzw. Muskelmasse kann ein gefiederter Muskel deutlich mehr Kraft entwickeln! Der Nachteil dieses muskelphysiologischen „Bautricks“ besteht jedoch in der Konsequenz der Goldenen Regel der Mechanik: Die höhere Kraftentwicklung wird durch einen geringeren Muskelkontraktionsweg „erkauft“, weswegen gefiederte Muskeln vor allem für kraftvolle, aber kleine Bewegungen geeignet sind. Im menschlichen Körper sind sie daher üblicherweise auch an genau den hierfür prädestinierten anatomischen Orten ausgeprägt. Spindelförmige Muskeln hingegen sind dort zu finden, wo üblicherweise ausladende, schnelle Bewegungen realisiert werden müssen. Wie der Muskelquerschnitt ist auch der Fiederungswinkel durch Training an die sportartspezifischen Erfordernisse in gewissen Grenzen anpassbar 5.
Berücksichtige, dass jeder Athlet sein persönliches muskuloskelettales Profil besitzt. Sowohl Hebellängen als auch das Muskelfaserspektrum sowie die Hypertrophieneigung bilden Bestandteile des sportartspezifischen biomechanisch-physiologischen Talents.
Vergegenwärtige dir die Anforderungen an die Arbeits- und Haltemuskulatur in deiner Sportart. Gestalte die Trainingspläne entsprechend.
- Ahonen, J. (1994). Sportmedizin und Trainingslehre. 1. Auflage. Nürnberg, New York: Schattauer.
- Badtke, G. (1999). Lehrbuch der Sportmedizin. Heidelberg, Leipzig: Barth.
- Herzog W. (2018). The multiple roles of titin in muscle contraction and force production. Biophysical Reviews, 10 (4), 1187–1199.
- OpenStax (2019). Zugriff am 16.05.2019 unter https://openstax.org/details/books/anatomy-and-physiology
- Alegre, L. M., Jiménez, F., Gonzalo-Orden, J. M., Martín-Acero, R. & Aguado, X. (2006). Effects of dynamic resistance training on fascicle length and isometric strength. Journal of Sports Sciences, 24 (5), 501-508.